
The Human Penis is Remarkably Boring
In case you missed it, I wrote an article for Quillette critiquing the first episode (titled ‘Monogamy, explained’) of Vox’s new Netflix series. Author Christopher Ryan was featured in that video, and he provided a very civil and thoughtful reply to my article in the comments section at Quillette.

He makes two main points here that I think are worth engaging with further. The second one (communal caregiving among foragers), I will save for a later date, to focus narrowly here on the first.
The first point is in regard to the suite of features that Ryan argues testify “in support of ancestral promiscuity.” These being: “penis morphology, repeated thrusting movement, frequent non-reproductive sexual behavior, female multiple orgasm, female copulatory vocalization, etc.).”
Let’s consider penis morphology. I will quote at length the case Ryan makes regarding the uniqueness of the human penis in Sex at Dawn:
Despite its lack of curlicues, the human penis is not without interesting design features. Primate sexuality expert Alan Dixson writes, “In primates which live in family groups consisting of an adult pair plus offspring [such as gibbons] the male usually has a small and relatively unspecialized penis.” Say what you will about the human penis, but it ain’t small or unspecialized. Reproductive biologist Roger Short (real name) writes, “The great size of the erect human penis, in marked contrast to that of the Great Apes, makes one wonder what particular evolutionary forces have been at work.” Geoffrey Miller just comes out and says it: “Adult male humans have the longest, thickest, and most flexible penises of any living primate.” So there.
Homo sapiens: the great ape with the great penis!
The unusual flared glans of the human penis forming the coronal ridge, combined with the repeated thrusting action characteristic of human intercourse—ranging anywhere from ten to five hundred thrusts per romantic interlude—creates a vacuum in the female’s reproductive tract. This vacuum pulls any previously deposited semen away from the ovum, thus aiding the sperm about to be sent into action. But wouldn’t this vacuum action also draw away a man’s own sperm? No, because upon ejaculation, the head of the penis shrinks in size before any loss of tumescence (stiffness) in the shaft, thus neutralizing the suction that might have pulled his own boys back. Very clever.
Intrepid researchers have demonstrated this process, known as semen displacement, using artificial semen made of cornstarch (the same recipe used to simulate exaggerated ejaculates in many pornographic films), latex vaginas, and artificial penises in a proper university laboratory setting. Professor Gordon G. Gallup and his team reported that more than 90 percent of the cornstarch mixture was displaced with just a single thrust of their lab penis. “We theorize that as a consequence of competition for paternity, human males evolved uniquely configured penises that function to displace semen from the vagina left by other males,” Gallup told BBC News Online. (Ryan, 234).
Broadly, we can break Ryan's argument down into a number of specific claims:
1) The human penis has “interesting [unique] design features,” it’s “specialized.”
2) The human penis is, among other virtues, the “longest…of any living primate.” (according to Geoffrey Miller, and quoted favorably by Ryan)
3) The flared glans of the human penis is “unusual” and that, according to Gordon G. Gallup, “as a consequence of competition for paternity, human males evolved uniquely configured penises that function to displace semen from the vagina left by other males.”
Now, Sex at Dawn was published in early 2010; however, in 2009, a book was published by primatologist Alan Dixson, who Ryan accurately describes as a “primate sexuality expert.” Ryan repeatedly quotes Dixson’s work from the late 1990’s and early 2000’s in Sex at Dawn, but I’m guessing Dixson’s 2009 book, Sexual Selection and the Origins of Human Mating Systems, was published too late for Ryan to integrate his insights from that work into his own, since it is not referenced.
This is unfortunate, because Dixson offers a pretty comprehensive breakdown for why each of these claims are likely mistaken.
First, the human penis actually seems to be comparatively quite dull in terms of “design features”. Here are illustrations from Dixson’s work on the penile morphology of various primates.

A-H are from primates which have polygynous mating systems, while I-N are from primates with multi-male/multi-female mating systems. As you can see, primates with polygynous mating systems have comparatively simple penises, much more similar to that of humans, compared to those of multi-male/multi-female mating primates.
To better compare primate penile complexity, Dixson created a rating system:
Ratings of penile complexity (length, distal complexity, size of baculum, and penile spines) show some consistent differences between these mating systems, as can be seen in Figure 3.22. Using a 5-point scale, each trait was rated for species representing forty-eight primate mate genera…Primates in which the females typically show multi-partner matings (multi-male/multi-female and dispersed mating systems) have significantly longer and distally more complex penes than representatives of polygynous or monogamous genera (Figure 3.22). They also tend to have longer bacula (when present) and larger penile spines, but these traits are more variable.

Regarding humans, “Human penile morphology is not exceptional when compared to that of the prosimians, monkeys, and apes. The overall rating for all four traits analysed is 10 for H. sapiens. This is the same rating as scored by a number of putatively monogamous or polygynous primates (e.g. Leontopithecus, Callimico, Erythrocebus, Theropithecus) and less than the ratings given to twenty-seven of the forty-eight primate genera included in the study.”
So in regard to penile morphology, humans are comfortably within the norm of monogamous and polygynous primates, and unlike multi-male/multi-female mating primates.
Next, let’s consider size. In Sex at Dawn, Ryan shows this graph:

The problem with this graph is that it compares erect human penises with flaccid bonobo, chimp, and gorilla penises. Dixson provides a table comparing erect and flaccid penis length between chimps, bonobos, gorillas, orangutans, and humans:
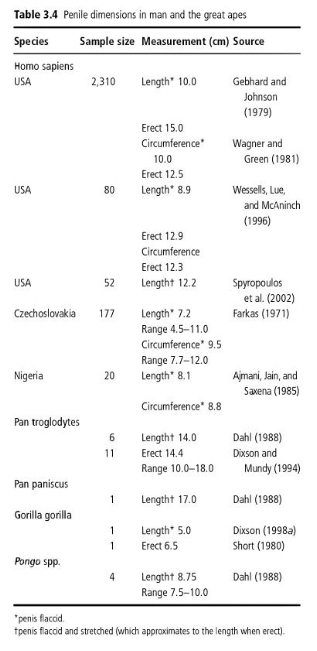
As you can see, when erect, human penis length is actually roughly the same as that of chimps and bonobos. Further, Dixson writes that, “The erect human penis is comparable in length to those of other primates, in relation to body size.”

So the human penis is not uniquely long: it is roughly the same length as that of chimps and bonobos, and it scales with body size to about the same degree as most other primates. Its size is not particularly impressive.
However, when considering its girth, the human penis might be at least somewhat unique. Dixson adds that, “Only its circumference is unusual when compared to the penes of other hominoids. However, even this comparison may not hold true for some primates, such as the spider monkeys (Ateles) which have large and thick penes.”
I can't find any comparative data on penis circumference, in Dixson's work or elsewhere, so this question has to be tabled. Dixson writes that, "No accurate data on penile circumferences are available for apes, or for the vast majority of primate species."
Nonetheless, even if the human penis is uniquely thick (which has yet to be demonstrated) there is no reason to think this has anything to do with sperm competition or "ancestral promiscuity".
Finally, what about Gallup’s ‘semen displacement’ hypothesis? Again, Dixson must disappoint. I will quote him at length here, but you can skip to the bolded parts to get the gist of his argument.
Gallup et al. (2003) tested Baker and Bellis's (1995) hypothesis concerning the 'plunger' action of the human glans penis in relation to sperm competition. They used models of human penes and vaginae to examine the putative effects of copulatory movements upon displacement of previously deposited (artificial) semen. Despite their contention that the large diameter of the human glans and its posterior margin (corona) represent adaptations to displace semen and provide an advantage in sperm competition, I can find no comparative evidence to support this view. A helmet or acorn-shaped glans is common amongst Old World monkeys, such as various colobines, macaques, baboons, mangabeys, and guenons, regardless of whether they have polygynous or multi-male/multi-female mating systems (see Figure 3.21 for examples). Gallup et al. stress that reduced length of the portion of the penile shaft covered by the prepuce (the pars intrapreputialis) in man is unusual, citing this as an adaptation to assist in removing semen from the vagina. However, this trait is shared by H. sapiens and the gorilla which, despite having a very small penis in relation to its body size, exhibits a distal morphology more similar to the human condition than is the case for other apes… The gorilla's genitalia are remarkably similar (in miniature) to the human condition. Among the African apes, the chimpanzee and the bonobo have the most specialized and derived penile morphologies. A glans penis is lacking, and distally the penis is filiform and contains a very small baculum (6.9 mm in P. troglodytes and 8.5 mm in P. paniscus)…The information presented thus far has, I hope, helped the reader to place earlier accounts of human penile morphology and that of apes and other primates in comparative perspective. A detailed argument has been made here because I believe it necessary to correct misunderstandings about the role of sexual selection and sperm competition in relation to the evolution of the human genitalia. It is highly unlikely that penile size or shape in human beings has been influenced by sexual selection via sperm competition…Nor is there any credible evidence that human penes evolved as plungers to displace semen deposited by previous males.
So the human glans penis is not actually unique, and is in fact very similar to that of numerous Old World Monkeys, and much of human penis morphology is actually quite similar (though larger) to that of gorillas (who have a polygynous mating system). In contrast, chimps and bonobos (with multi-male/multi-female mating systems) have a much more specialized penile morphology, and yet they lack the glans penis that Gallup (likely incorrectly) considers to be an adaptation for sperm competition (which is extreme among chimps and bonobos).
I’m afraid your dick is remarkably boring.
Now, I haven’t delved deeply in regards to the reasons for “frequent non-reproductive sexual behavior, female multiple orgasm, female copulatory vocalization, etc,” in humans, but I’m going to go out on a limb here and propose that for any argument that associates these behaviors with “ancestral promiscuity,” you could equally come up with a model that considers them to be adaptations for pair-bonding in humans. Or by-products of other features. I’ll look a bit more into it, though, and If I find anything notable I’ll do a follow up.
Otherwise, in part 2, I’ll address Ryan’s claims in Sex at Dawn, and in his reply to my article, about communal caregiving among foragers.